 IV. Forskjeller i hvordan vårt DNA blir benyttet -Del II
IV. Forskjeller i hvordan vårt DNA blir benyttet -Del II
(fra 'Theistic Evolution - a Scientific, Philosophical and Theological critique, kap.15 -- slutten)
Når det gjelder hva som gjør oss menneskelige, er det ikke hva som er likt, men hva som er ulikt som teller. J. Marks (1)
Bilde 1. Relevant bok i så måte
Oppsummering:
Overvekten av bevisene sier at vi mennesker har et unikt opphav. I boka 'Theistic Evolution' (A scientific, philosophical and theological ) vises at neo-darwinismen ikke kan forklare det informasjonsbærende komplekset, og spesifiserte ved livets natur. Vi skal her se at den heller ikke kan gjøre rede for spesifikke mønstre vi kan se i vårt genom og at felles avstamning for sjimpanser og mennesker er tvilsomt. Mengden av og type forskjeller i vårt DNA kan ikke gjøres rede for i den foreslåtte evolusjonære tidstabellen. De tilsynelatende hensiktsfulle plasseringer av repeterende elementer som SINES og LINES, samt deres viktige roller i artsspesifikke ulikheter, argumenterer mot en ikke-styrt prosess av felles avstamning. Den rene forbløffende kompleksitet i genregulering og kromosom interaksjon, argumenterer for briljant design. Våre anatomiske og fysiologiske forskjeller ville kreve mange spesifiserte, koordinerte endringer, som ikke kunne inntreffe uten styring. Men viktigere, om ting som delt kodon-benyttelse og pseudogener kan forklares funksjonelt (vs tilfeldig), smuldrer argumentet for felles avstamning bort, og en unik opprinnelse blir mer sannsynlig.
 IV. Forskjeller i hvordan vårt DNA blir benyttet
IV. Forskjeller i hvordan vårt DNA blir benyttet
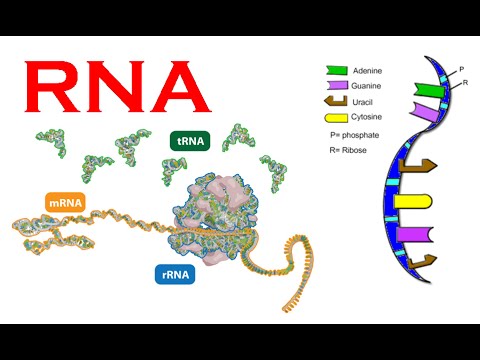
Bilde 2. RNA kan benyttes på finurlige måter
Tilbake til 1970-tallet foreslo Mary-Claire King og A.C. Wilson at forskjellene som betyr mest kunne være i hvordan DNA ble benyttet. (10) Vi vet nå at hvor, når og hvordan DNA blir benyttet betyr mye mer enn et antall nukleotide-forskjeller. Menneske-spesifikke forskjeller i genregulering, er det som gjør oss unike.
Menneske-spesifikke gener.
Vi har i overkant av 20 tusen gener, og vi vet nå at noen er unike for oss. Estimater på hvor mange varierer fra 300 til 600 spesifikke protein-kodende gener, som kan skyldes duplikater av eksisterende gener. (11,12) Disse genene synes ofte å være uttrykt i hjernen eller testiklene.(13) Så mange som 60 nye (de novo) gener er også rapportert. Disse genene synes å ha en effekt, f.eks. i forhold til sykdom. I den sammenheng er det verdt å merke seg at endog et nytt gen er en bemerkelsesverdig ting fra et evolusjonært synspunkt. Det var akseptert at det å få nye gener, syntes meget vanskelig, men nå blir nye gener funnet i hvert genom som blir sekvensiert, gener som synes å være unike for rasen (orfan-gener). Men om det var overraskende for evolusjonære biologer, så passer det godt i et design-syn på livet.
Flerfunksjons gener
I slutten av 1970-årene ble det oppdaget at protein-kodende gener kan bli benyttet til å kode for mange ulike proteiner. Gener deles opp i segmenter (eksoner), og etter genene er transkribert til RNA, kan RNA bli spleiset i ulike arrangement av eksoner. Om et gen har eksonene 1-5, kan RNA bli spleiset slik at ekson 1,2,3.4, eller 1,3,4 evt. 4,5. Det kan også være gener som uttrykkes i motsatt retning, f.eks. 5,3,2. Så selv om vi kun har drøyt 20 tusen gener, så har vi potensielt hundre-tusenvis av ulike protein-kodende RNA som kommer fra disse genene, der hvert unikt RNA lager ett ulikt protein. (13) Det er en "viktig rolle for alternativ spleising i å etablere forskjeller mellom sjimpanser og mennesker" (16) Faktisk 6-8% som har blitt studert, viser spleise-forskjeller mellom sjimpanser og mennesker. Dette innebærer at våre gener kan benyttes til å lage ulike proteiner, selv om de synes å ha samme DNA-sekvens.
Ulike gen-uttrykk
Ulike vev og organer i kroppen lages av ulike celle-typer, og hver celle uttrykkes sitt eget sett av gener. Uttrykks-mønsteret for gener avgjør hvilke celletyper de blir. (17) For at vi skal vokse til å bli mennesker, og ikke sjimpanser, er det sannsynlig at forskjeller i genuttrykk er involvert. Signifikant er det substansielle forskjeller i hvordan gener uttrykkes, spesielt i hjernen. (18) Det er ulike slag forskjeller involvert i å regulere ulike typer av gen-uttrykk. Den mest velkjente er protein-transkripsjons faktorer (hvorav 1-3 % er menneske-spesifikke). (19) Transkripsjons-faktorene gjenkjenner og binder seg til spesifikke bindings-steder for DNA, og influerer aktiviteten i omkringliggende gener. Noen øker, noen minker, gen-uttrykk. Noen er vevs-spesifikke, og noen er generelle forsterkere av gen-uttrykk. Noen stilner gener fullstendig. 1-3% av våre transkripsjons-faktorer er unike for oss mennesker. En enkelt transkripsjons-faktor kan endre uttrykket av mange gener, og har således en sterkt forsterkende effekt. Disse unike transkripsjons-faktorene kan ha en signifikant effekt på hvordan vårt genom benyttes, selv om de selv utgjør en meget liten del av den totale DNA-mengden.
Ikke-kodende DNA og dets funksjoner
Bare omtrent 2% av vårt genom bærer informasjonen som lager protein, et forbløffende faktum. (20) Ca. halvparten av det ikke-proteinkodende DNA er sammensatt av repeterende elementer, noen ganger kalt mobile genetiske elementer. Mange av disse repeterende elementene ligner et visst slag av viralt DNA, som kan kopiere seg selv og sette seg inn i nye lokasjoner i genomet, selv om mesteparten av disse mobile elementene er inaktive. I mange år gikk mange vitenskapsfolk ut fra at ikke-proteinkodende DNA var søppel-DNA. Men vi vet nå at noe av dette 'søppel'-DNA er involvert i å modulere oppførsel og struktur i kromosomer, som regulerer gen-uttrykk, og kanskje å forsyne måter organismer kan respondere på sine omgivelser genetisk.
Fr. Collins uttrykker dette skifte i synet på'søppel-DNA', slik det rapporteres i World magazine: "I termer om 'junk-DNA' så benytter vi ikke lenger den termen, fordi jeg tror det var mye lik en hybris å forestille oss at vi kunne kvitte oss med noen del av genomet, som om vi visste nok til å si at det ikke var funksjonelt.. Mesteparten av genomet som vi trodde var der for å 'fylle ut', viser seg å ha funksjon." (For ordens skyld gir ikke Collins opp forestillingen om at mennesker og sjimpanser har felles stamfar.)
Vi skal nå se på ulike slag 'junk-DNA' fordi de utgjør en stor del av vårt genom, og har i mange tilfeller vist seg å ha funksjon.
SINES (Short Intersperced Nuclear Elements) er en slags mobilt genetisk element som utgjør ca. 12% av genomet. Bl.a. hjelper SINES til å spesifisere i hvilke celler gener skal uttrykkes. Sju tusen Alus (en slags SINE) er arts-spesifikke, tilstede i mennesker, ikke sjimpanser.(23) SINES synes å finnes i nærheten av gener av spesielle funksjonelle typer, og påvirke deres uttrykk, -ofte i spesifikke celle-type måter. (26) Menneske-spesifikke Alu-elementer er også virksomme i RNA-editering, en prosess som endrer RNA, etter den er kopiert fra DNA. (27) RNA-editering kan påvirke mange aspekter av RNA-prosessering, slik som spleising og stabilitet. (28) Slik SINE-indusert RNA-editering er mest bemerkelsesverdig i den menneskelige hjernen, og er i hovedsak arts-spesifikk. (29) Selvsagt er hjernen et av de områder vi skiller oss mest fra sjimpanser. Men RNA-editering skjer ikke bare i hjernen. Den er essensiell for utvikling og feilaktig editering resulterer i sykdommer som kreft og psykiske lidelser. (30)
LINES er et annet mobilt genetisk element, kalt (Long Intersperced Nuclear Elements). LINES utgjør ca. 17% av genomet. Mange (ca. 1.800) er artsspesifikke. (31) De har en måte å regulere måter kromosomet oppfører seg, hvor de knytter seg til kjernen, hvordan de pakkes, og forhindrer at gener uttrykkes. (32) De spiller slik en sentral rolle i organisering av cellekjernen. De tenderer også å flankere grupper av gener, som for å kontrollere utrykk av RNA fra ulike posisjoner i DNAet. De ser ut til å tegne konturene av gen-uttrykk. (33) LINEs hjelper også styre utvikling av ulike celletyper i hjernen. LINEs flytter til nye lokasjoner i genomet til spesielle hjerneceller i det de utvikler seg, og havner nær ulike gener involvert i neuronal utvikling. Det innebærer at mange av vår hjernes celler skriver om sin genetiske instruksjon i løpet av prosessen til å bli modne neuroner. (35) Det samme er tilfelle også i annet kroppsvev. Det skulle være tydelig at slike arts-spesifikke elementer (SINEs og LINEs) kan utgjøre dyptgående forskjeller i vår anatomi, fysiologi og oppførsel. Spesielt i våre hjerner.
Lange ikke-proteinkodende RNA (lncRNA)
 70-90% av genomet er transkribert til RNA, men produserer ikke protein. Slike lange ikke-proteinkodende RNA kan være nøstet innen gener, i begge retninger, eller de kan komme fra DNA med ikke-proteinkodende gener. (36) Ofte stammer de fra, eller inneholder, SINEs og/eller LINEs. Slike lncRNA synes å være arts-spesifikke og har ofte funksjon, når de undersøkes. De virker gjerne til å endre DNA-arkitektur. Noen linker ulike DNA-strekk sammen, looper DNA til funksjonelle domener, eller forårsaker endring i gen-uttrykk. Andre knytter DNA til kjerne-periferien når de ikke trenges. Andre igjen er buffere for å hindre at gen-uttrykk fra ulike DNA-segmenenter i DNA fra å blande seg med hverandre, (38) hvilket i så fall kan medføre kreft. Mange av dem er vevs-spesifikke. F.eks. finnes det menneske-spesifikke lncRNA uttrykt i hjernen. (40)
70-90% av genomet er transkribert til RNA, men produserer ikke protein. Slike lange ikke-proteinkodende RNA kan være nøstet innen gener, i begge retninger, eller de kan komme fra DNA med ikke-proteinkodende gener. (36) Ofte stammer de fra, eller inneholder, SINEs og/eller LINEs. Slike lncRNA synes å være arts-spesifikke og har ofte funksjon, når de undersøkes. De virker gjerne til å endre DNA-arkitektur. Noen linker ulike DNA-strekk sammen, looper DNA til funksjonelle domener, eller forårsaker endring i gen-uttrykk. Andre knytter DNA til kjerne-periferien når de ikke trenges. Andre igjen er buffere for å hindre at gen-uttrykk fra ulike DNA-segmenenter i DNA fra å blande seg med hverandre, (38) hvilket i så fall kan medføre kreft. Mange av dem er vevs-spesifikke. F.eks. finnes det menneske-spesifikke lncRNA uttrykt i hjernen. (40)
HAR. (Human Accelerated Regions) er menneskelige DNA-segmenter hvis segmenter skiller seg substansielt fra relaterte pattedyr-sekvenser. Pattedyrsekvensene er 'konserverte' --de skiller seg lite arter i mellom. Men menneskelige HAR-er helt annerledes, hvilket kan skyldes at de skyldes design. Slike HAR-er har en tendens til å være lokalisert nær 'utviklings gener' transkripsjons-faktorer og gener uttrykt i sentral-nervesystemet. (41)
Organisering av våre DNA-sekvenser skiller seg betydelig fra hverandre. Vi har ulike rekombinatoriske hot-spots i forhold til sjimpanser. Disse hot-spotsene er plassert hvor homologe kromosomer rekombinerer, ved å skyfle DNA-segmenter inn i nye kombinasjoner. Som følge er vårt DNA organisert i ulike 'skyflede' blokker. Innsettinger på mellom 10-15 tusen DNA-baser utgjør samlet 32 millioner baser av menneske-spesifikt DNA. Innsettinger på mer enn 15.000 baser utgjør ca. 8 millioner baser av menneske-spesifikt DNA. Endelig skiller vårt kromosom 2 seg fra sjimpansers (se slutten av kapitlet).
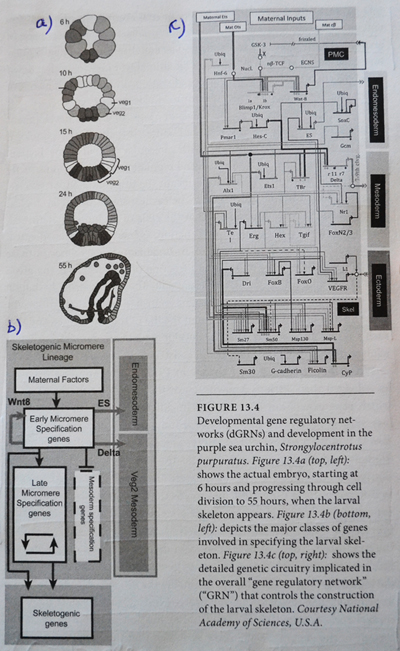
Bilde 5. dGRN hos sjøpinnsvin
Genetiske nettverk (dGRNs)
Transkripsjonsfaktorer, SINEs, LINEs og lnc-RNA virker alle til å regulere gener i et slags genetisk interaksjons-nettverk. Innen dette nettverket fungerer noen gener som forbindelses-punkter -der endring kan påvirke mange andre gener, som samhandler over nettverket. (44) Således kan endring av gen-uttrykk for noen få gener, ha stor virkning. Det er estimert at ca. 17% av det nevrale nettverk i hjernebarken er unike for menneske, jamvel om genomet totalt sett bare kan skille seg 5% fra sjimpanser. (45) Andre deler av hjernen viser også andre forskjeller. Det innebærer at små endringer i gen-regulerende elementer, kan ha store virkninger på våre gen-nettverk, og således på våre fysiske og oppførsels-karakteristika.
Genomet er et operativsystem. OM genomet virker komplisert, er det fordi det trenger å være det. Det trengs for å aktivere timing og lokalisering av ulike gen-uttrykk. Gener grupperes i mapper og supermapper. DNA organiseres til looper, så mega-mapper; kromosomer har sine strukturer i kjernen, hver med sitt eget territorium, slik at settet av gener kan bli uttrykt på en koordinert måte. Gener i aktive mapper behandler ulike input, og samordner i en samordnet output. De fungerer lik logiske sluser. Utdata til alle de logiske cellulære gater kombineres, og resultatet er nøye regulert cellulær oppførsel, og til slutt et fungerende menneskelig vesen. Det er hvordan DNA benyttes, ikke hvor mye som er felles, som avgjør forskjellen mellom mennesker og sjimpanser. (46)
V. Fysiologiske og anatomiske forskjeller
Gitt de genetiske og regulatoriske kompleksitetene ovenfor, er det ingen overraskelse at vår fysiologi atskiller seg den hos sjimpanser. Vi har ikke samme reproduserende biologi, tennene våre utvikler seg senere etter fødselen, våre barn fødes helt hjelpeløse og krever forlenget pleie av mødre. Våre hjerner er større og fortsetter å utvikle seg etter fødselen, og former neuron forbindelser i et rasende tempo. Vår muskulatur er svakere, med mindre innsettingspunkter. Vårt hormonsystem, immunforsvar, diett/fordøyelse, svømmeevne, evne til å gråte skiller seg fra sjimpanser. våre føtter er designet for å gå rett opp og ned, mens sjimpansers for å klatre i trær. Våre nakker er lengre, hodeskallen er plassert på toppen av ryggraden for balanse og kunne se framover. Vår brystkasse kan beveges fritt fra side til side for å veie opp for vår gangart, samt at den utvider seg for å tillate dyp pusting mens vi løper. (48) Våre skuldre er designet for å kaste, mens sjimpansens for å klatre. (49) etc., etc., etc.
En artikkel kalt "Comparing the Human and Chimpanzee Genomes: Searches for Needles in a Haystack," av Arjit varki og T. Altheide, beskriver bokstavelig mer enn hundre fysiologiske og atferdsforskjeller. Bramble og  Lieberman, lister i journalen Nature endringer som er nødvendige for lang-distanse løping, inkludert 26 anatomiske og fysiologiske innovasjoner. (54) Det mest slående trekket som skiller oss fra sjimpansene er vårt intellekt. Det er ingen fullgod evolusjonær forklaring for dets oppdukking. Varki et al. hevder: "Det er vanskelig å forklare hvordan konvensjonell naturlig seleksjon kunne ha utvalgt på forhånd for de forunderlige evner i det menneskelige intellekt, som vi fortsetter å undersøke i våre dager. .. Forklaringer basert på 'eksaptasjon' (tilpasning av trekk opprinnelig beregnet for noe annet), synes utilstrekkelige. Mesteparten av det det menneskelige sinn gjør i dag, eksistere ikke engang, da bevisstheten opprinnelig utviklet seg. Eksperter i menneskelig evolusjon og erkjennelse har enda til gode å forsyne en skikkelig tilfredsstillende forklaring." (55)
Lieberman, lister i journalen Nature endringer som er nødvendige for lang-distanse løping, inkludert 26 anatomiske og fysiologiske innovasjoner. (54) Det mest slående trekket som skiller oss fra sjimpansene er vårt intellekt. Det er ingen fullgod evolusjonær forklaring for dets oppdukking. Varki et al. hevder: "Det er vanskelig å forklare hvordan konvensjonell naturlig seleksjon kunne ha utvalgt på forhånd for de forunderlige evner i det menneskelige intellekt, som vi fortsetter å undersøke i våre dager. .. Forklaringer basert på 'eksaptasjon' (tilpasning av trekk opprinnelig beregnet for noe annet), synes utilstrekkelige. Mesteparten av det det menneskelige sinn gjør i dag, eksistere ikke engang, da bevisstheten opprinnelig utviklet seg. Eksperter i menneskelig evolusjon og erkjennelse har enda til gode å forsyne en skikkelig tilfredsstillende forklaring." (55)
Bilde 6. En hjerne -- to bevisstheter
En kan selvsagt finne spekulasjoner om dette på Internett og i museer. Favoritt-gener listes som årsaks-agenter for utvikling fra ape-lignende stamfedre. Men for at felles avstamning fra disse skulle være sann, måtte mange av disse trekkene oppstå samtidig, på koordinert vis. Det innebærer at et antall mutasjoner måtte skje samtidig. F.eks. ville en forstørret hjerne kreve endringer til kjeven og tennene så vel som til hodeskallen, noe som ville kreve at diverse mutasjoner inntraff noenlunde samtidig. En artikkel i Ass. Press diskuterte et scenario der en enkelt genetisk mutasjon, funnet i mennesker, ble foreslått å ha redusert størrelsen til våre forfedres kjeve-muskulatur, og tillate større hodeskalle og hjerne å utvikle seg. Kritikere er uenige, og sier at evolusjon ikke fungerer så greit: "De første mennesker med mutasjonen ville trolig hatt veikere munn, men likevel hatt store tenner og kjever. Mange mutasjoner i tillegg ville ha vært nødvendige. "Mutasjonene ville ha redusert darwinistisk fitness til individene," sier antropolog B. Wood ved G. Washington Universitet. "De ville bare ha blitt fikserte om den sammentraff med mutasjoner som reduserte tannstørrelse, kjeve-størrelse og økte hjerne-størrelse. Hva er sjansene for det?" (56)
 Slike mutasjoner kunne ikke har vært hva slags mutasjoner som helst, eller feil rekkefølge, ellers kunne de ha vært ødeleggende eller dødelige. Som sagt måtte mange mutasjoner ha inntruffet samtidig for å fullføre den type koordinert endring som er nødvendig for vår evolusjon. Men gir historien tilstrekkelig tid for det?
Slike mutasjoner kunne ikke har vært hva slags mutasjoner som helst, eller feil rekkefølge, ellers kunne de ha vært ødeleggende eller dødelige. Som sagt måtte mange mutasjoner ha inntruffet samtidig for å fullføre den type koordinert endring som er nødvendig for vår evolusjon. Men gir historien tilstrekkelig tid for det?
VI. Tid nok for endring? -henviser til denne linken her.
Bilde 7. Enorm komprimering av DNA
VIII. Argumenter brukt til støtte for felles avstamning
Pseudogener. Fordi pseudogener synes å lage defekt protein, evt. ikke noe protein, har disse vært ansett som 'søppel', levninger fra en tidligere evolusjonær prosess. De blir tatt som bevis for felles avstamning fordi de er plassert på samme sted og synes å ha de samme 'feil' i mennesker og sjimpanse (Fr. Collins og D. Alexander). Som nevnt er synet på 'søppel-DNA' drastisk endret. (64) Deler av problemet er at pseudogener kan være aktive i visse vev bare i visse deler av veksten, noe som gjør identifikasjon av deres virkemåte vanskelig. Forskere i feltet er visse på at videre forskning vil frembringe mer bevis på funksjonalitet. En gruppe uttalte: "Vi tror at flere og flere funksjonelle pseudogener vil bli oppdaget i det nye biologiske teknologier vil bli utviklet i framtiden .. så avgjort, de såkalte pseudogenene er virkelig funksjonelle og flere gjenstår å bli oppdaget og utforsket i diverse organismer." (65) Det er flere måter pseudogener er funnet å virke på så langt. De kan fusjonere med nabogener for å produsere enten produserende eller ikke-produserende RNA, eller de kan danne RNA-RNA duplekser med sitt 'foreldre'-gen og forhindre uttrykking eller signalisere dets nedbryting. (66)
Ett tegn som peker i retning av sannsynlig funksjonalitet er at deres sekvens er meget like i flere ulike arter. Det finnes mer enn 8 tusen pseudogener i det menneskelige genom; 60% er like for mennesker og mus. Det kan bety at de har en sekvens-avhengig essensiell funksjon, slik at sekvensen ikke kan endres uten skade. (67) Den grad av likhet er ikke noe som kunne forventes om pseudogener ikke tjente noen biologisk funksjon. Om de har en funksjon i både menneske og sjimpanser som er avhengig av presise sekvenser, så ville ikke likheten mellom menneske og sjimpanse være underlig. Som ett eks. viser pseudogener i beta-globin ansamling av gener hos mennesker likhet innen og mellom menneske og ape- populasjoner. Men denne likheten skyldes ikke protein-koding, men synes å involvere regulering av utviklingsendringer via kromosom-interaksjoner. (68)
Synteny refererer til hvor godt kromosom-sekvenser fra ulike arter matcher hverandre. Hvis de matcher godt, er dette tatt som tegn på en felles stamfar. Men det finnes en mulig funksjonell forklaring: Kromosom-struktur har en dyptvirkende effekt på gen-regulering og hvor gener er lokalisert i kjernen påvirker hvordan de uttrykkes. (69) Genomer er høyt ordnede, likevel dynamiske enheter der kromosom posisjoner, strukturer og interaksjoner er kontrollert for å regulere kjerne-prosesser i cellen. (70) Dette raffinementet er et sterkt argument for design. Således er det mulig at likhet i genorden kan være en funksjon både mennesker og sjimpanser trenger. Dersom behovet er funksjonalitet, trenger det ikke indikere felles avstamning.
 Synonymt kodon-bruk. Ett av 'bevisene' for felles avstamning, er 'gjenbruket' av kodoner. Noen slike tripletter av nukleotid-sekvenser i DNA (kodoner) spesifiserer samme aminosyre, f.eks. lysin. Lysin kan være kodet via fire ulike kodoner i RNA (CUU, CUC, CUA eller CUG). Formodningen har vært at disse kodonene er synonyme, og burde kunne brukes om hverandre. Likevel, når det er valg mellom ulike kodoner, benytter sjimpanse og menneske alltid det samme kodonet. Det er i senere tid oppdaget at kodoner er benyttet for flere formål, og hvilke kodoner som benyttes påvirker DNA-sekvensen, som igjen kan påvirke gen-uttrykket. (71) Protein-degradering (kontrollert ødeleggelse av proteiner i cellen) eller hvordan proteiner folder seg til 3-dimensjonale former. (72) Kodoner kan derfor være identiske fordi de må utføre multiple jobber, som ingen av de alternativene (for samme aminosyre) kan utføre. Dette kan vel så gjerne være grunnen til at sjimpanser og mennesker benytter samme kodon, som felles avstamning :-) Dette er også et sterkt argument for design: Det sofistikerte ved å ha koder innenfor koder, ville ikke oppstå ved enkle mutasjoner og naturlig seleksjon.
Synonymt kodon-bruk. Ett av 'bevisene' for felles avstamning, er 'gjenbruket' av kodoner. Noen slike tripletter av nukleotid-sekvenser i DNA (kodoner) spesifiserer samme aminosyre, f.eks. lysin. Lysin kan være kodet via fire ulike kodoner i RNA (CUU, CUC, CUA eller CUG). Formodningen har vært at disse kodonene er synonyme, og burde kunne brukes om hverandre. Likevel, når det er valg mellom ulike kodoner, benytter sjimpanse og menneske alltid det samme kodonet. Det er i senere tid oppdaget at kodoner er benyttet for flere formål, og hvilke kodoner som benyttes påvirker DNA-sekvensen, som igjen kan påvirke gen-uttrykket. (71) Protein-degradering (kontrollert ødeleggelse av proteiner i cellen) eller hvordan proteiner folder seg til 3-dimensjonale former. (72) Kodoner kan derfor være identiske fordi de må utføre multiple jobber, som ingen av de alternativene (for samme aminosyre) kan utføre. Dette kan vel så gjerne være grunnen til at sjimpanser og mennesker benytter samme kodon, som felles avstamning :-) Dette er også et sterkt argument for design: Det sofistikerte ved å ha koder innenfor koder, ville ikke oppstå ved enkle mutasjoner og naturlig seleksjon.
Bilde 8. Histon-kode for lesing av oppkveilet DNA
Kromosom-funksjoner. Kromosom 2 hos mennesket synes å være en fusjon av to kromosomer hos sjimpanse. Dette har vært fremført som et sterkt argument for felles avstamning. Men stedet der fusjonen fant sted, består ikke av typiske telomer-sekvenser (som finnes i enden av kromosomene). I stedet er det funnet degenererte sekvenser, som er funnet andre steder i genomet, men ikke er assosiert med brudd eller fusjoner. (73) Det menneskelige kromosom 2 kan alltid ha vært som det framtrer nå. Det er ikke noen spesiell grunn til å foreslå noen fusjon, uten under formodning av felles avstamning. Dermed kan det ikke benyttes som et argument for felles avstamning. (74) {Ville i så fall forutsette det en ønsket å vise -tigge spørsmål' -oversetters tilføyelse.}
Argumentet om designeren som en luring, går på hvorfor han fikk det til å se ut som ting utviklet seg (makro-evolusjon)? Men at ting ser ut som de er utviklet, er på bakgrunn av evolusjonsteorien (ET). Biologer blir kontinuerlig fortalt at de må huske at 'ting bare ser designet ut, de er det ikke i virkeligheten'. (76) Det innebærer at designeren ikke er en luring'. Han har gjort det slik for at alle kan oppdage hans design. (Rom1v19-20).
 IX. Konklusjoner
IX. Konklusjoner
Den overlegne bevisbyrden sier at vi mennesker har et unikt opphav. Den tilsynelatende hensiktsmessige plassering av repeterende elementer som SINEs og LINEs, samt deres viktige rolle arts-spesifikke forskjeller, argumenterer mot en ikke-styrt prosess av felles avstamning. Den skjære kompleksiteten i gen-regulering og kromosom interaksjon argumenterer for et briljant design. Våre anatomiske og fysiologiske forskjeller ville kreve mange spesifikke, koordinerte endringer, som ikke kunne skje uten styring. Enn videre, dersom ting som synteny, delt kodon bruk og pseudogener kan bli forklart funksjonelt, så forvitrer argumentet om felles avstamning, og en unik opprinnelse blir mer sannsynlig.
Det sentrale spørsmålet om menneskelig evolusjon er dette: stammer vi fra ape-liknende stamfar, eller er vi unike med et distinkt opphav? En av grunnene til at folk blir teistiske evolusjonister er at de betrakter bevis for genetiske likheter med en ape-liknende stamfar for å være uomtvistelige. Dette kapitlet viser at disse bevis ikke er så ugjendrivelige når det kommer til stykket. Forskjellene mellom menneske og ape er mye større enn det som allment er blitt rapportert. Genetiske forskjeller viser mange funksjonelt betydelige, unikt menneskelige trekk. Det er for mange slike endringer til å ha skjedd ved tilfeldige mutasjoner og naturlig seleksjon. I tillegg viser det som tidligere ble betraktet som 'søppel-DNA', å inneholde mange regulerende elementer, hvorav noen er artsspesifikke. Selv om de ikke tallmessig er store, har de en betydelig rolle i hvordan genomet vårt fungerer, spesielt i hjernen. På bakgrunn av manglende kjennskap til mye av genomets virkemåte og i lys av økende bevis for spesifikt menneskelig design, så er det en dårlig idé å skrive om teologien (for å passe til evolusjonsteorien).
Bilde 9. Junk-DNA motbevist
Referanser:
35. K.R. Upton, D.J.Ferhardt et al, "Ubiquitous L1 Mosaicism in Hippocampal Neuron," Cell 161 (2015): 228-239
36. Mercer et al, "Long Non-coding RNAs"
38. Johnny T.Y. Kung et al, "Long Non-coding RNAs. Past, Present, and Future," Genetics 193 (2013): 651-669
40. Perdomo-Sebogal, "Gene Regulation Factors."
41. M.J. Hubisch et al, "Exploring the Genesis and Functions of Human Accelerated Region Shed Light on Their Role in Human Evolution," Current Opinion in Genetics and Development 29 (2014): 15-21.
 44. M. Fontenot et al, "Molecular Networks and the Evolution of Human Cognitive Specializations," ," Current Opinion in Genetics and Development 29 (2014): 52-59
44. M. Fontenot et al, "Molecular Networks and the Evolution of Human Cognitive Specializations," ," Current Opinion in Genetics and Development 29 (2014): 52-59
45. Oldham et al, "Conservation and Evolution of Gene Coexpression Networks in Human and Chimpanzee Brains, " 17973-17978.
46. R. Sternberg and J. A. Shaphiro, "How Repeated Retroelements Format Genome Function," Cytogenetic and Genome Research, 110 (2005): 108-116.
48. D. Bramble et al, "Endurance Running and the Evolution of Homo," Nature 432 (2004): 345-352
49. N. T. Roach et al, "Elastic Energy Storage in the Shoulder and the Evolution of High-Speed Throwing in Homo," Nature 498 (203): 483-487
50. Varki and Altheide, "Comparing the Human and Chimpanzee Genome."
54. D. Bramble and Lieberman, "Endurance Running
55. Vaski et al, "Explaining Human Uniqueness."
56. J.B. Verrengia, "Scientists Debate Suggestion Jaw Mutation Led to Evolution," Ass. Press. USA Today, March 24, 2004.
64. Balakierev and Ayala, "Pseudogenes: Are they 'Junk'?"
65. Y. Z. Wen et al., "Pseudogenes Are Not Pseudo Any More," RNA BIology 9, no.1 (Jan. 2012): 27-32
66. D. Zheng and M.B. Gerstein, "The Ambigious Boundary between Genes and Pesudogenes: The Dead Rise Up, or Do They?" Trends in Genetics 23 (2007): 219-224.
67. Y. Z. Wen et al., "Pseudogenes Are Not Pseudo Any More,"
68.A. Molerinho et al, "Evolutionary Constraints in the beta-Globin Cluster: The Signature of Puryfying Selection at the delta-GLobin (HBD) Locus and Its Role in Dev. Gene Regulation," Genome Biology and Evolution 5 (2013):559-571
69. St. A. Hoang et al, "The Network Architecture of the Sacharromyces cerevsiae Genome," Plos One 8 (2013): e81972
70. C.D.M. Rodley et al, "Global Identification of Yeast Chromosome Interactions Using Genome Confor mation Capture," Fungal Genetics and Biology 46 (2009): 879-886
71. A.B. Stergasis et al, "Exonic Transcription Factor Binding Directs Codon Choice And Affects Protein Evolution," Science 342 (2013):1367-1372
72. F. Zhang et al, "Differential Arginylation of Actin Isoforms Is Regulated by coding Sequence-Dependent Degradiation,2 Science 329 (2010):1534-1537.
73. Y. Fan et al, "Genomic Structure and Evolution of the Ancestral Chromosome Fusion Site in 2q13-2q14.1 and Paralogous Regions on Other Human Chromosomes," Genome Research 12 (2002):1651-1662.
74. A. Gauger et al, Science and Human Origins (Seattle, Discovery Institute Press, 2012)
76. R. Dawkins, The God Delusion (New York: Random House, 2009).
Oversettelse og bilder ved Asbjørn E. Lund